Articles - Israel's Corner
The articles presented here are some contemplations and research by the webmaster. We hope that you find them interesting and useful.
Are Cages Evil? - A look at keeping birds in captivity
By Israel
The hobby of keeping birds will invariably involve some sort of cage or enclosure which can range in size from small to massive. It is a necessity in fact, but there will always be those who are against it, thinking it morally wrong to keep an animal in captivity, that it is cruel to strip them of their freedom. They might even liken it to battery hens versus free range hens, though the only similarity there is that they are both birds. There is such a thing as a cage too small (especially in the aforementioned case), but no cage at all can also be more cruel. Some time ago an activist went to a bird show, and in protest opened the cages of as many birds as she good, letting them fly free. And condemning them to death, as they could not survive in the wild.

|
| Yay, fresh greens :) |
There are many aspects, viewpoints and factors to cages and the keeping of birds in captivity, and all things considered the birds of many hobbyists have a much more enjoyable life than those out in the wild - even to the extent that they will try and join their ranks!
Many of the birds kept as pets and bred in New Zealand cannot survive in the wild here, as our country does not have the correct environment for them, be it lack of appropriate food sources or breeding areas, the severity of the weather patterns, or the existence of predators that would be quite happy to have them as a snack. Included in this number are budgies and canaries. Raw NZ is just not suitable.
Many birds like security and stability - for them, the cage is a barrier that keeps the outside world out, not them in. It's a place that they call home, to feed and breed and be themselves without having to worry about being upset or disturbed. This can quite easily be demonstrated by the birds that, when let out of their cages, fly around for a while before going back to the comfort of their cage by themselves. For them it is their home.
Captive birds, that are well looked after and cared for, never have to worry about food at all. Every day, no matter what the weather or season, there is plentiful seed, greens and whatever else available for them. Enough to go around too! This is attractive enough to some wild birds that they will break in to aviaries, make themselves at home, and then refuse to leave, even breaking in again if removed! This behaviour has been seen particularly with Sparrows, but can also be clearly seen with birds that locate a stable food source and stick around, such as with plazas in towns, supermarkets, public gardens, fishing vessels and so forth.
A result of captivity over time (or close association with humans such as birds in public places) is that the birds become domesticated (to varying levels dependant on species), and loose some of their wild instincts. This leaves them unable to survive by themselves, and though it is caused by our actions in the first place it is difficult to reverse.

|
| An indoor flight cage. |
Size in important though! There is such a things as too small, though minimum cage size does depend on purpose and breed. Aviaries are obviously the best and come in many different sizes and designs, and are capable of housing many different varieties, though some types of birds should not be mixed. The larger the aviary the more flight space for the birds, and the more birds that can be kept. For outdoor aviaries precautions do need to be taken to prevent disease and predators (which are often more lethal to captive birds if they manage to get past the defences). Inside, aviaries are not always practical, so the next best thing is flight cages. They are much larger than the typical cages that you see in pet stores and many homes, and allow the birds to fly around and exercise, as well as allowing a small number of birds to be housed together. For birds, length is often more important than height, as they get exercise from flying horizontally and not vertically. There are 'standard' cages of a large enough size, but they are only useful for keeping the smaller birds, and even then flights are a much better option. Inside, birds have less threat from predators (except perhaps the household cat), but care does need to be taken that they experience the natural day cycle, for their benefit. Inside is invariably warmer though. As a note, breeding cages are sized for practical purposes, to enable mating pairs to have the best chances at breeding and rearing their young, and similarly for show cages (they are temporary - it is not their permanent home).
In conclusion, there are both benefits and drawbacks to keeping birds in captivity. When done properly, with due care and affection (or practical considerations) life is much easier for the birds, albeit different from what they would experience in their native lands. Yes, they have lost their freedom (generations breed in captivity may not even known what that is), but the return for them is significant, and some even seek that lifestyle.
The keeping of birds is an enriching experience and hobby, and can bring a lot of joy and fulfilment to the lives of both the keeper and kept. The world would be a duller place without it. From this author there is no regret :) [And his birds enjoy a comfortable life inside in flight cages]
Copyright Israel 2012. Used with permission.
Record Keeping
By Israel
So, you're going to breed birds. Throw a pair together, add in some nesting material. That's all you need to do right?
Well, not quite. While that may work in some cases (especially for colony breeding where you just leave the whole lot of them to do their own things), there are many factors & elements that need to be considered, especially if you are trying to breed for particular traits or the elusive show winner. In order to be able to make well informed decisions it is important to keep records - information on your birds.

|
| Budgie Record Sheets on breeding cages. |
As to how to store the information, where and what to record, there are many options. There are basics that are essentially requirements to good record keeping and breeding, but the general rule of thumb is the more you record about each bird the better. The better you know your birds, the more success you will have.
So why keep records?
First and foremost, you won't remember everything about all your birds. Even if you start out small, eventually you may end up with a lot of birds, possibly in the hundreds (some of the bird club members have had 600+ birds at one time), and by that point it is easy to lose track or forget things.
If you are pairing birds with particular traits in mind, then knowing the ancestry of both of the birds is vital. Genetics is somewhat complicated, with some traits being dominant, recessive, sex-linked, single and double factors, as well as traits being able to be carried but not present. What you see isn't always what you have, and sometimes the resulting chicks can be surprising. Some traits can be so dominating that if not paired correctly all you get is that trait. A little knowledge of particular bird genetics and good records can go a long way to help predicting what the offspring are likely to carry.
Similarly, if you are line breeding it is essential to know which birds are related and how closely, as well as who the original source birds came from. Also, if deformities arise in your birds, you'll want to know who the parents were, and what other birds may be carrying that gene unseen.
Some birds are good parents, dedicating themselves to their young, while others are not, fluttering from nest to seed dish, oblivious to their duties. Some males are able to fill all of the hens eggs, while others barely fill one. Some hens build awesome nests, while other may find their vulnerable young to be a good source of nesting material, stripping them of their feathers. Some males may tirelessly feed their young while other may decide that they are in the way and want to start a new clutch. All of these characteristics are important considerations when deciding on matching, and even whether or not to keep the birds in the long term. Some birds are good for breeding, others simply are not. But even so, if you decide to use the not so good, awareness of what they might do is useful so that you can foster out the young, or sub in a foster parent to help out so that losses are minimised. Some of these traits can also be inherited by the young, so knowing what habits you are passing on is useful.
Age is another factor. There is a minimum age before they can successfully start breeding, and they can also get too old. Birds of a similar age usually breed better together as well.
When buying birds, you'll usually have a purpose in mind, and records will help you know what birds you got from where, and how you integrated them into your breeding lines. Similarly, when culling or selling birds, records will aid you in the decision making process of what birds to keep and what ones to get rid of. Ancestry and habits are important traits fro birds, as well as how they physically look.
|
| Canary records in a text document. |
So what needs to be recorded?
Anything that you think might be useful later!
Essential information to record includes (info that should be also included on breeding cards is in italics):
- form of identification for the birds (be it name, band details, description)
- gender (once learnt)
- date hatched
- parents
Optional information includes (but is by no means limited to):
- date paired, and who with
- breeding box numbers/details (birds often breed better in the cages (or same environment) that they were born in)
- number of eggs laid, number fertile, number hatched, number of clutches etc
- transfers, fostering etc
- parenting skills
- visual/split traits/genes
- feather type
- behavioural traits
- health traits & issues had
- status (alive/dead/sold/loaned etc)
- date last treated/wormed
- who the bird was brought from or sold to (and for how much), and if the bird is marked for someone
- shows entered and results
- date the bird passed away
- ...and anything else you might think useful.
|
| Budgie records software (The Budgerigar Program). Very in-depth. |
Finally, how should this information be recorded and kept?
Most breeders use a double system - simple breeding cards for use on the cages that contain the essential information that is needed and gathered at that point, and software or a more comprehensive record on paper/pc for more permanent and in-depth information. There is a variety of (somewhat expensive) bird and animal breeding software on the internet, enough to suit most people, and though they require a bit more effort, creating a tabulated database in a book or on a PC isn't too difficult.
Remember, though it may seem like lots of effort, all information you keep will be invaluable, and will enrich your enjoyment of (& success in) the hobby!
Many thanks to Lynn & Mike who contributed information to this article. Copyright Israel 2012. Used with permission.
The Genetics of Colour Canaries
A look at mutations in Colour Canaries, and pairing expectations.
By Israel
Genetics. While it may seem like a daunting topic, a basic understanding of genetics can prove to be helpful to breeders, aiding in the decisions when pairing two birds, knowing what will and won't work, what could give rise to complications, and helping determine expectations of what the offspring might be like.
|
| Genetics, wikipeda style. |
Genetics is the study of genes, heredity and variation. All of which are of interest to breeders. Genetic data is stored in DNA, and a gene is a collection of DNA that control some hereditary trait. A chromosome is an organised structure of genes, and they always come in pairs. Different species have different numbers of pairs of chromosomes, and the order in which genes are stored on the chromosomes, and the difference in the number, determines whether or not cross-breeding is viable between the species. Human have 23 pairs, while canaries have 21. When mating occurs, half of the genetic material comes from the father and the other half comes from the mother, and this is what gives rise to genetic variation in species.
In order to be able to study how genes are passed from parent to child we will be using Mendelain Genetics. Gregor Mendel was a Austrian priest/monk, who studied trait inheritance in garden peas and wrote a paper on it. The importance of his work was not realized until well after his death when other rediscovered his principles. Mendel summarized his findings in two laws: The Law of Segregation, and the Law of Independent Assortment. The Law of Segregation states that every individual possesses a pair of genes for a given trait, and that the parent randomly passes one of these genes to its offspring. (so one comes from one parent, and one from the other). The Law of Individual Assortment states that separate genes for separate traits are passed along to the offspring independently of each other. These laws are represented using Punnet Squares.
Traits have different 'types' for determining how they interact with each other. The two simplest ones are Dominant and Recessive. If a trait is dominant then it will show if it is present in either of the genes present in the pair, while if it is recessive it will only how up if it is the only trait present in the pair.
Let's use the colour of a flower as an example. We'll classify red as dominant (R, dominant genes are always represented by upper-case), and white as recessive (r, recessive is always lower-case). Therefore, a red flower, as it is the dominant colour would be represented either by RR (homozygous, with the dominant trait on both genes) or Rr (heterozygous, dominant recessive mix). In the latter case the flower is split for white (or carries white), as it has the gene, it just doesn't show. The white flower would be repesented by rr.
Traits aren't always simple though, and there are other possibilities as well. A trait can be semi-dominant, resulting in a blending between the two options, or co-dominant, in which case both options display themselves. In the flower example above, if red and white were semi-dominant then the colour for a heterozygous flower would be pink, while if red and white were co-dominant then a heterozygous flower would have both white and red flower petals. Finally a trait can be dominant (lethal), where a homozygous combination of the trait is non-viable. In the example above RR would be lethal, resulting in the death of the flower at an early stage in development, and so if a flower was red it would have to be Rr (a non-lethal combination).
Traits can also be sex-linked, which all things considered aren't too difficult to deal with, except when you start considering crossovers. Sex-linked traits are, as their name suggests, linked to the sex genes, which are a special set of genes. In humans, and most species, the male is heterozygous and represented by XY, while the female is homozygous, represented by XX, and the father determines the sex of the child. In birds and butterflies however, the male is homozygous (ZZ), while the female is heterozygous (ZW). Sex-linked traits are always recessive, and are carried on the Z gene, so for example the agate mutation would be represented by Za. As hens only have the one Z gene, if they possess the trait it shows, while males can be split/carriers for the trait, as they have to have it on both genes for it to show.
Now that we have all that out of the way, it's time to look at some examples. Punnet Squares are used to examine the expectations of a pairing. It is traditional to put the male along the top and the hen on the side.

|
| Yellow variegated Gloster Corona |
For our first example let's look at Gloster Canaries. There are two types of Gloster canaries, Coronas and Consorts. Coronas have the crest, consorts lack it. When pairing two glosters together it is recommended that coronas are paired with consorts, and this is because the crest gene is dominant (lethal). Let us label the crest gene as C and the lack as c; CC is a non-viable combination of the gene as it is lethal and kills the chick early in development, so a corona would be represented by Cc, while the consort would be represented by cc.
So if we did pair two coronas together the punnet square would look like this:
| C | c | |
| C | CC | Cc |
| c | Cc | cc |
There are four possible ways that the genes can combine following Mendels laws, giving three results. Based on the number of times they turn up in the square we can assign them a percentage expectation, but this expectation is a per chick basis, and not a nest average. Each chick has the same probabilities of having a particular set of genes, and over a large number of chicks you would expect it to average out, but on a small scale it often may not.
So for this combination the expectation is:- 25% change of being CC - non-viable combination
- 50% chance of being Cc - corona
- 25% chance of being cc - consort
If we went with the recommended pairing of corona to consort:
| C | c | |
| c | Cc | cc |
| c | Cc | cc |
The expectations this time are:
- 50% chance of being Cc - corona
- 50% chance of being cc - consort
No dead chicks this time around!
Glosters are a type canary, but the crest gene can be present on any colour of canary.
For our second example lets have a look at a the agate mutation, which is a sex-linked (recessive) trait, and we'll pair a split male (ZZa) with an agate hen (ZaW).
| Z | Za | |
| Za | ZZa | ZaZa |
| W | ZW | ZaW |
The expectations from this pairing is:
- 25% split male (ZZa) - remember as the trait is recessive it won't show if not present on both genes.
- 25% agate male (ZaZa)
- 25% normal hen (ZW)
- 25% agate hen (ZaW)
If we had paired a agate male with a normal hen:
| Za | Za | |
| Z | ZZa | ZZa |
| W | ZaW | ZaW |
The expectations would then be:
- 50% split males (ZZa)
- 50% agate hens (ZaW)
Now onto the more interesting stuff - mutations and colouring.
There are two 'layers' to the colouring of canaries. There is the base (ground) colour, also known as the Lipochrome (after the fat that dyes the feathers), and on top of that is the dark brown and black pigmentation of the feathers (Melanin). When this pigmentation is broken and light feathers of the ground colour show then the canary is considered to be variegated. If there are no dark feathers at all, and the canary is just showing the ground colour then the canary is said to be Clear.
Canaries have three base colours - Yellow, White, and Red. Variegation comes in two layers - black and brown, and a canary can be anything from completely variegated, to clear of variegation, and anything in between. Variegation and colour are separate, and there are mutations that effect both of these.
The wild Green canary is a yellow based bird, with unbroken melanin (in other words 100% variegation; in this state the bird is considered Self). This is the dominant state for canaries, and self canaries of any of the three base colours (without any other mutations effecting their colouring) are considered the Black/Natural Series. The appearance of variegation in captive canaries, as well as clear canaries is caused by multiple recessive mutations, and the interaction of these genes is what determines how much variegation the bird has (and hence isn't quite as simple as the examples thus far given).
The following tables (for those set-up the same as the one below) list different names/types and series for the various combinations of colour and mutations in canaries.
Ground Colours & Variegation:| description // ground colour | Yellow** | White* | Red** |
| Black Series | |||
| All dark, with no light feathers (Self) melanin is black & dark brown (and all other colours): | Green | Blue / Slate | Bronze |
| Partially variegated: | Yellow variegated | Silver | Red variegated |
| Clear (no variegation - still possess melanin though): | Yellow (Lipochrome) |
Dominant White* Recessive White* |
Red (Lipochrome) |
** Both yellow and red come in different shades, and this can effect the name which is reflective of the shade. There are at least two other factors that effect the shade, a saturation gene, and feather type. More on both of these later.
Levels of variegation are classified as follows:
| Self | - All dark, with no light feathers (unbroken) |
| Foul | - One or two light feathers in wing flights or tail |
| Heavily Variegated | - More than 75% but less than 100% dark markings |
| Medium Variegated | - Between 50% - 74% dark markings |
| Lightly Variegated | - Less than 50% dark markings |
| Ticked | - Only 1 dark marking smaller than a dime |
| Clear | - No variegation at all |
Note that for different types of canaries the definition for light to heavy variegation can vary.
So how do the various levels of variegation interact? As variegation is controlled by multiple genes that are not independent, their behaviour cannot be easily modelled using a punnet square, but experience from centuries of breeding has given an indication of what can be expected in general terms, splitting canaries into the three groups of melanin (self), variegated and clear. As a note, the Black, Brown, Isabel & Agate series are all collectively termed as Melanin birds (and you can get them as self, and as variegated).
So, the expectations when pairing melanin (self), clear and variegated are as follows:- melanin x melanin = 100% melanin
- melanin x clear = 100% variegated
- melanin x variegated = 50% melanin, 50% variegated
- variegated x variegated = 25% melanin, 50% variegated, 25% clear
- variegated x clear = 50% variegated, 50% clear
- clear x clear = 100% clear
Of course this does not provide any indication of the level of variegation that one could get on a particular bird, and for the purposes of this table ticked is grouped with clear, and foul with self. A note on modelling - the gene that produced variegation is recessive, and the gene responsible for clear birds is also recessive (and could be the same one perhaps), but looking at the above expectations it behaves more like a semi-dominant trait when fully combined, and can be modelled as such at the most basic level. To do this we would use V and v, VV being self/foul, Vv being variegated and vv being clear/ticked. Of course, as for the above expectations list, it cannot show exactly how much variegation to expect.
There are two forms of white - Dominant White and Recessive White. Dominant whites can have a faint yellow tinge on the curve of the wing and on the outer edges of the primary flight feathers (as the mutation 'pushes' the yellow colour into the background, but not 100% completely), while recessives do not. Dominant white is also dominant (lethal), and so a doubling up of the gene is non-viable. However, this does make it slightly easier to deal with from an expectation point of view. As a point of clarification this does effect all mutations/series based on the colour, so a Blue bird is still a white based bird, and hence still has the lethal factor.
Dominant white is dominant to both red and yellow, while recessive white is recessive to both red and yellow, which is fairly straight forward. Red and yellow are both semi-dominant with respect to each other, which is not so simple. This dominance can be represented by the following flow:
Dominant White > Yellow = Red > Recessive White
Let's have a look at the colour expectations. We'll pair a dominant white (split for yellow, so we'll represent it by Yy) with a yellow (yy). Remember that YY is non-viable.
| Y | y | |
| y | Yy | yy |
| y | Yy | yy |
The expectations are:
- 50% dominant white (Yy)
- 50% yellow (yy)
If we paired dominant white to dominant white, then as for the gloster example above we would expect 25% chance of being non-viable (YY), 50% chance of being white, and 25% chance of being yellow.
Now let's look at recessive white (xx), pairing with a yellow split for white (Xx):
| X | x | |
| x | Xx | xx |
| x | Xx | xx |
This time round the expectation is 50% chance of being recessives white, and 50% chance of being yellow, split for recessive white.
Yellow and red are not so simple, and there are a number of factors involved for both.

|
|
Feather types: (Left): Frosted/Buff; (Right): Intense |
The first is feather type (which effects all colours/types, and not just canaries either). Canaries have two feather types, Intensive and Non-intensive. Intensive (yellow, non-frosted) feathers are narrower, harder, and the colour goes right to the edge of the feather. This gives the canaries colour a more intense appearance. Non-intensive (buff, frosted) feathers on the other hand are broader, softer, with white tips, and soften the colour some what. Sources seem to indicate that Intense is a dominant (lethal) trait (so an intense bird would be represented by Ff), and so buff would recessive (ff). Continual buff to buff breeding can give rise to very soft feathers that have trouble coming out through the skin and end up causing feather lumps, while intense to intense breeding can lead to very narrow feathers that don't fully cover the bird (this hints that the trait isn't quite as simple as a plain dominant lethal trait, but it can still be modelled under that assumption). The recommendation is that pairing should always be Intense to Non-Intense, and this gives 50% intense, 50% non-intense.
The second is the colour saturation factor, which is modelled as a semi-dominant trait. Splitting yellow into three shades, light, medium and dark, light yellow is absent the trait (ss), medium yellow has it once (Ss), and dark yellow has it twice - a double factor (SS). Combining these two together gives rise to six distinct shades, which is of course not exhaustive, and both of the above apply to the red ground colour also.
Feather Type and colour saturation can be modelled using punnet squares.
Finally, both red & yellow are semi-dominant. When the cross-breeding was first attempted to create a red canary by pairing a red-siskin finch with a yellow canary, the assumption was that the red gene would push out the yellow as they both lie on the same position on the chromosomes, but given their nature this was not the case. Instead we ended up with orange canaries. The assumption is that there are numerous genes (instead of one) that control the colour red, but simple modelling is still possible.

|
|
Heavily variegated orange-bronze Canary chick. |
Taking into consideration just the third factor (for simplicity, and the results are still useable), let's have a look at some pairings.
Pairing an orange (RY, the mid-way point between red and yellow) with an orange:
| R | Y | |
| R | RR | RY |
| Y | RY | YY |
The expectation is:
- 25% red (RR)
- 50% orange (RY)
- 25% yellow (YY)
Pairing red to yellow would be 100% orange, while pairing yellow to orange would be 50% yellow, 50% orange, and red to orange would be 50% red and 50% orange.

|
| Non-colour fed red factor canary. |
[As an aside, canaries have the ability to convert yellow lutein into the yellow colouring for their feathers. Red-siskins have the ability to convert yellow lutein into the red colour. The cross breeding did successfully pass on this ability to the canaries, but instead of having just one, they now possess both. Yellow lutein is present in carotene, and so is a natural way of colouring the feathers (for both red and yellow), but many of the other red-food supplements dye the feathers directly by-passing this process. Of course the feathers can only go red if they have the ability to do so, otherwise using the red dye just ends up causing splotches on the feathers.]
Back to mutations:
The following is a mutation that effects the lipochrome colour:| description // ground colour | Yellow** | White* | Red** |
| Ivory / Lipochrome-Pastel | |||
| Reduction of the strength of the lipochrome colour to a pastel like colour. Sex-linked recessive: | (Gold) Ivory | Silver Ivory | Rose Ivory |
The following mutations all effect the melanin and are Sex-Linked Recessive (and have an associated dominance flow):
| description // ground colour | Yellow** | White* | Red** |
| Agate (series) | |||
| Dilutes of the Black Series , the black (& brown) melanin is reduced to grey. | Gold-Agate | Silver Agate | Rose Agate |
| Brown (series) | |||
| Only brown melanin present (Cinnamon factor#) - black is either suppressed or converted. | Cinnamon (Gold-Brown) | Fawn (Silver-Brown) | Brown-Red? |
| Isabel Series^ | |||
| Dilutes of the Brown Series, the brown melanin is of a sandy colour | Gold Isabel | Silver Isabel | Red Isabel |
| (Melanin)-Pastel^^ | |||
| Reduction of the strength of the melanin colour to half strength. | Gold Pastel | Silver Pastel | Rose Pastel |
| Suppressed black melanin, and almost suppressed brown melanin (on top of feather and quills only), Red eyes. %% | Satinette % | ||
| Visual difference between the sexes - sex-linked, dimorphic and ??? | Mosaic |
% Satinette carriers can be visually discerned for agate series.
%% Exists in all four series (Black, Agate, Brown & Isabel), and comes out slightly differently depending on the series. Agate carries show that they are split for the trait, and assumed that crossovers have an impact on it too.
^ Results from a crossover (see below)
^^ crossover is necessary for manifestation in agate and black series, and can effect all of the series it is recessive to.
As you will have noticed there are a number of different mutations that effect the melanin of canaries, and their effects can be mutually exclusive (but not always). Hence, as mentioned in the table label, they have an order of dominance:
Black > Agate = Brown > Isabel > Melanin-Pastel = Satinette.

|
|
Satinette Canary chick. Her red eyes don't show up so well in this photo. |
So, what happens when you cross two sex-linked mutations? Lets illustrate with an example. Let's take a brown series hen (ZbW) and pair her with an agate male (ZaZa):
| Za | Za | |
| Zb | ZaZb | ZaZb |
| W | ZaW | ZaW |
The expectation is:
- 50% Agate Hens (ZaW)
- 50% Green Male I (ZaZb) - he appears as a normal green male, however he is split for both agate and brown! (he is green as he does not have the required double factor for either trait in order for them to be expressed)
[Notation note - type I males are split for two traits, while type II males are expressing one sex-linked recessive trait and still split for two others! ]
During Meiosis (a special type of cell division that produces sperm and eggs) the chromosomes split apart, so that the child gets one from each parent. For homologous chromosomes (chromosomes of approximately the same length such as the Z chromosome), there is the possibility of a crossover, where the chromosome breaks (they are fragile after all) and then recombines with its matching pair. It is not as common as the normal gene split but it does occur, and with a male who is split for two different sex-linked traits it opens up an interesting possibility (and this is the main area where its occurrence can be noticed). In the case of the example we are currently looking at it gives rise to Isabels.
So, if we take the Green male I from above and pair him with a brown hen, the standard expectations would be:
| Za | Zb | |
| Zb | ZaZb | ZbZb |
| W | ZaW | ZbW |
- 25% green male I (ZaZb)
- 25% brown male (ZbZb)
- 25% agate hen (ZaW)
- 25% brown hen (ZbW)
This is of course a much different result than what a normal green male would have produced.
If a crossover occurs when the genes are splitting, then instead of getting the expected ZaZb --> Za & Zb from the above male, we would end up with ZaZb --> Zab & Z.
The expectations from the pairing with a crossover occurring are:
| Z | Zab | |
| Zb | ZZb | ZabZb |
| W | ZW | ZabW |
- 25% Green male split for brown (ZZb)
- 25% Green female (ZW)
- 25% brown male II (split for isabel and agate) (ZabZb)
- 25% Isabel hen (ZabW)
As the agate mutation reduces the intensity of the black melanin, the assumption here is that when it is paired with the brown mutation on the same gene it will act to reduce its intensity instead (black has been suppressed/converted into brown already), hence becoming the Isabel factor. The two individual components can still work as intended to express brown/agate if correctly paired as well1.
Following on, if we pair the brown male II with an brown hen:
| Zab | Zb | |
| Zb | ZabZb | ZbZb |
| W | ZabW | ZbW |
- 25% brown male II (as above; ZabZb)
- 25% brown male (ZbZb)
- 25% brown hen (ZbW)
- 25% Isabel hen (ZabW)
And for the final example in this section, if we pair an isabel hen with a brown male II:
| Zab | Zb | |
| Zab | ZabZab | ZabZb |
| W | ZabW | ZbW |
- 25% Isabel male (ZabZab)
- 25% brown male II (ZabZb)
- 25% Isabel hen (ZabW)
- 25% brown hen (ZbW)
The Ino mutation is a recessive mutation that also effects the melanin of the birds. Unlike the mutations above it is not sex-linked.
| description // ground colour | Yellow** | White* | Red** |
| Red eyes, Suppressed black melanin, brown melanin on flight feathers on edges only. | Ino |

|
|
The satinette canary from above (now 6 months), and her fawn sibling. |
The Phaeo, Opal and Onyx mutations are also all recessive (but not sex-linked), but as they are not very common in NZ they won't be covered in any depth in this article at this stage.
The final area to look at is the expectations for multiple traits at once. Punnet squares can be used to model more than one trait at once, but the number of rows/columns doubles for each trait being looked at. One trait has two rows/columns, two traits has four, three has eight and so forth. Hence one trait has 4 possible outcomes, two have 16, three have 64 and so on. We will look at some two trait punnet squares, three traits and more however are beyond the scope of this article (space constraints).
The simplest pair of traits to look at (that aren't two sex-linked genes; as can be seen above they are able to be modelled on a small punnet square) is a sex-linked recessive trait (we'll take the cinnamon factor) and a dominant/recessive colour match-up (white/yellow). Let's look at the pairing of a cinnamon yellow hen (ZbWyy) with a dominant white male split for fawn and yellow (ZzbYy):
For each bird there are four combinations of the genes that can be given to the offspring.
| ZY | Zy | ZbY | Zby | |
| Zby | ZZbYy | ZZbyy | ZbZbYy | ZbZbyy |
| Zby | ZZbYy | ZZbyy | ZbZbYy | ZbZbyy |
| Wy | ZWYy | ZWyy | ZbWYy | ZbWyy |
| Wy | ZWYy | ZWyy | ZbWYy | ZbWyy |
Notice however that two of the four gene combinations for the hen are identical to two others, resulting in two rows being duplicated. For simplicity and ease of reading they could be removed from the table.
The resulting expectations are:- 12.5% White (split for fawn) Male
- 12.5% Yellow (split for cinnamon) Male
- 12.5% Fawn White Male
- 12.5% Cinnamon Yellow Male
- 12.5% White Female
- 12.5% Yellow Female
- 12.5% Fawn Female
- 12.5% Cinnamon Hen
- 25% White (split for fawn) males
- 25% Yellow (split for cinnamon) males
- 25% White Hens
- 25% Yellow hens.
For another example, let's pair a cinnamon yellow hen (ZbWYY) with a red-factor male (ZZRR). The combinations of genes for just these two traits are fairly simple - the males combination is only ZR (the full four being ZR, ZR, ZR, ZR), and the hen can contribute either Zby or WY (the full four being ZbY, ZbY, WY and WY). Given that simplicity, let's try adding the modelling of variegation to the mix with V representing self/melanin, v representing clear, and both states modelled as semi-dominant. If the male is lightly variegated his notation would now be ZZRRVv, giving him two possible combinations (ZRV and ZRv), and the hens notation, if she was self, would be ZbWYYVV, still only giving two combinations (ZbYV and WYV).
| ZRV | ZRv | |
| ZbYV | ZZbRYVV | ZZbRYVv |
| WYV | ZWRYVV | ZWRYVv |
So the expectation would be:
- 25% orange-base black series male split for brown
- 25% orange-base variegated male split for brown
- 25% orange-base black series hen
- 25% orange-base variegated hen
For our final example lets try nutting out a real life example, throwing in a little detective work and a bunch of assumptions. A yellow isabel hen was paired with a fawn male. Out of four chicks there was two isabel yellows (one male, one female), one fawn (male), and a satinette (gender unknown at this stage). Ancestry for the parents in unknown, but as the satinette is a sex-linked recessive trait, the male has to be split for it, and the chick female. As we have both a male and a female yellow isabel, the male must be split for it as well. (There are at least two other options, but given the results thus far let's go with the simplest approach). Finally assume that the white ground colour for the fawn is Dominant white, and that the satinette is brown series. So the fathers notation would be ZbsZabYy, while the mother is ZabWyy. In the chicks we are looking for ZbsWYy (the satinette), ZabWyy and ZabZabyy (yellow isabels) and ZbsZbYy (fawn male, who could also be a carrier ZbsZabYy). Two lines of the punnet square have been removed as they are duplicated.
| ZbsY | Zbsy | ZabY | Zaby | |
| Zaby | ZabZbsYy | ZabZbsyy | ZabZabYy | ZabZabyy |
| Wy | ZbsWYy | ZbsWyy | ZabWYy | ZabWyy |
Using the above approach and assumptions all the chicks have been accounted for. Accuracy over a larger pool of examples would validate the model used for these birds. Crossovers would also be worth considering, especially if the fawn male chick was found to not be split for satinette or the satinette turns out to be isabel; the split would be Zb & Zabs instead of Zbs & Zab.
As with all things, the more information you have the better the results. Expectation modelling for coloured canaries is no different. Accurate records over many generations goes a long way with helping to make informed decisions when pairing canaries, especially if trying to breed to particular traits. Hopefully the information found within this article will prove useful to some and enrichen the enjoyment of this rewarding hobby.
Appendixes:
Article is currently version 2.33
List of notations used for genes in canaries in this article, and thoughts on ones that could be used:| ZZ | male bird |
| ZW | female bird |
| C / c | Gloster Crest (& lack of) |
|
Y / R y / r |
dominant white to semi dominant yellow / red |
|
x / t X / T |
recessive white to semi-dominant yellow / red |
| Y / R | semi-dominant yellow to red |
| Za | agate series gene |
| Zb | brown series gene (cinnamon factor) |
| Zi | ivory |
| Zab | isabel |
| Zp | pastel |
| Zs | satinette (most common is brown satinette Zbs) |
| I / i | (lack of) Ino |
| F / f | intense / buff |
| S / s | saturation factor |
| V / v | variegation (modelled as semi-dominant) |
The Red Canary, Tim Birkhead, 2003
Canaries and related birds, Horst Bielfld, 1981
The Practical Canary Handbook, Marie Miley-Russell, 2005
The Canary Handbook, Matthew M Vriends
Coloured Canaries, G.B.R Walker, 1976
Fundamentals of Genetics in Colour Canaries, Octavio Perez-Beato, 2008
http://www.aviculturalsocietynsw.org/GeneticsI.htm
1 This assumption is correct, and the notation used in this article works although it does not represent the actual genes involved. The usual notation used is Z BNO for a green canary, where B is the brown melanin, N/n is the presence/lack of the black melanin, and O/o is the oxidisation (dilute) factor/lack there of. So an agate is therefore BNo, a cinnamon is BnO and an isabel is Bno.
Copyright Israel 2013. Used with permission.
Mutants
A contemplation of deformities in caged birds
By Israel
Finding a deformed hatchling in one of our nests this morning came as quite a surprise to us. It was our first this season, first ever in fact, very unexpected, and somewhat heartbreaking as we mulled over our options of what we should do with it and what its chances of survival were. We talked to another breeder about it, and we hummed and hurred for a portion of the day, eventually reaching a decision that we were happy with. Next time the decision will likely be much quicker, but we would hope there isn't a next time, and having checked the rest of the brood we were relieved to find that it was a single occurrence.
But it got me to thinking on the topic of individuals born with deformities, and the why, how and what of it all. For many breeders it will be an occurrence that they are used to, for others it will be a rarity, and for some lucky ones it will be something that only happens to others.
A deformity occurs when something goes wrong somewhere - a gene becomes mutated during the formation of the chick, or a complication occurs during the development in the egg or later. The mutation can be as small as a change in plumage (which is how we got some of the varieties of our birds!) or a misformed toe, to things like dual beaks, crooked legs or even three legs. Severer deformities can occur, but they get to a certain point where the chick becomes non-viable and dies in development. I guess you could also consider some mental problems like an unwillingness to breed under the same umbrella as they have some of the same causes, though the manifestation is quite different.
So why do instabilities in the genetic make-up of our birds occur? Radiation is one possible cause, known to cause mutations in genes and outside of our control. There is much more radiation present in our world now than there used to be, and to see the results of an overdose at an extreme level, one need look no further than the tragic results of a nuclear power plant meltdown and the after effects. Cross-breeding is another possible cause, though not something that most breeders involve themselves in unless they are trying for mules or hybrids. The chromosomes between two species aren't always 100% compatible, and the combining of them can have unforeseen consequences. When successful though, mules turn out to be hardy and long-lived birds, just mostly infertile. The third cause is due to small gene pools, and there are three levels of this. First off, there are a limited number of birds in New Zealand and we are unable to get more in. This means that the gene-pool is closed and without cross-breeding we won't be able to add any variety into it (and cross-breeding can only do so much given that most offspring are infertile). For most varieties this isn't too much of an issue yet, but for some of the varieties with a smaller number the impact is becoming quite prominent, with breeding becoming more slow and difficult and an increase in the occurrence of deformities. The second is selective breeding. When we continually breed towards a particular trait or traits we are reinforcing certain genes, often at the expense of variety and other genes. The extreme version of this, which is the third level, is line breeding, where one breeds birds of the same lineage together, in order to again improve upon a trait. The gene pool become very small, and the chance of deformities increases significantly. But to some the consequences are worth it for the shows, and some varieties would disappear if it were not done.

|
|
This canary chick has a deformed left leg, and its feathers haven't properly grown on the left side either. However it is able to balance well on its good leg and fly. |
As to the what to do about it, there are only two options. Kill it or keep it. There are pros and cons to both options (more for one than the other), but compassion plays a part as well. In many industries deformed young are killed off pretty quickly or in some cases kept as pets, for example lambs, but the former is much more common. There are reasons for both options. Deformed birds are no good for breeding, as they have a significant chance of passing along the mutated gene, and hence they end up using resources for no return, and depending on the complication, they may also require extra care. They are likely to have shortened lifespans due to their unstable genes, surviving perhaps only a few weeks, sometimes much longer (a mule with three legs survived for nine months before dying). It is also a possibility that they will be picked upon by their cage mates, a part of the 'natural selection' process which could push them to an early grave, or that their condition could cause them continual discomfort, proneness to injury or difficulty with basic living. These are all reasons to end its life at an early stage. Attachment and compassion are the main driving factors for keeping them, to give them the best life that they could have (although some may see it as cruelty to prolong their doomed existence), to offer them something they would not have in the wild. And in some cases, you just can't bring yourself to do the deed. Either option has their repercussions and results, and in the end it is the decision of the individual breeder, as to what they feel is best.
Of course prevention is better than cure, but factors are not always under your control. If you don't know the ancestry of some of your birds then you are playing with unknown factors, and decisions made when pairing birds will effect the possibility of deformities. Record keeping as always is an invaluable tool, and will help in avoiding mistakes when pairing. But even still they can crop up. It's an aspect of the hobby, albeit a sad one, and we each will have to make our own decisions when they do.
Copyright Israel 2013. Used with permission.
Line Breeding
By Israel
In a recent bird club meeting the topic of line breeding was brought up, as well as the concept of breeders having their own 'lines' of birds. But what exactly is line-breeding, and how does one end up with their own line?
Line breeding is a type of breeding plan where birds of the same descent (related birds) are mated together over successive generations. This is done to try and maintain the characteristics and traits of good birds over successive generations or to make 'copies' of particular birds. It should be noted that you have to have good birds to start with as you are breeding from a closed gene pool. Because it is a closed gene pool being used, the good traits are hopefully reinforced, but on the other side of the coin any defects are also more likely to be reinforced and show as well. Hence careful selection and culling of birds is required in order to maintain the good quality. Note that line breeding is in-breeding, with a particular purpose in mind.
When line breeding the usual pairings used are father - daughter, and mother - son when trying to 'copy' a bird. If you are just trying to maintain or reinforce traits then projenitor - offspring, auntie - nephew, uncle - niece, half siblings, siblings, and possibly cousins (depending on how well they express the traits you are after) are also all pairing options, conditional of course on the quality of the birds.
Lets look at a simple example of how it works:
If we take two birds and pair them together, then the offspring (F1) are 50% their father, 50% their mother.
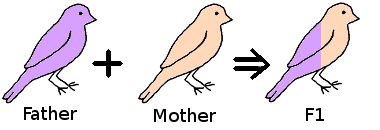
If we take a hen from the F1 generation and mate it back to her father then the resulting offspring (F2) is now 75% her father and 25% the original mother.
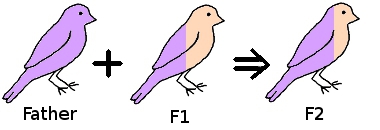
Taking a hen from the F2 generation, and mating her back to her father, the resulting offspring (F3) will be 87.5% their father, 12.5% the original mother.
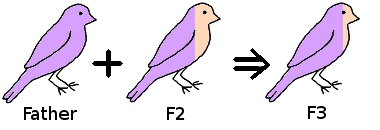
As can be seen the resulting chicks are becoming progressively more like their father and less like the original mother.
One of the possible outcomes from line breeding (apart from a closed gene pool) that breeders should be aware of is in-breeding depression, where fertility levels become low, some hatchlings end up being non-viable, some birds can end up being sterile, and other reproductive issues start appearing. In varieties that have had a small gene pool in New Zealand these issues are starting to show. This can often be solved by out-breeding (or cross-breeding) for a generation to a non-related bird, or one that isn't as closely related (such as a cousin), but this will of course add unknown or unwanted traits into the line.
The other main breeding technique is out-breeding, where two unrelated birds are bred together. The results from such pairings are not as predictable unless you have good ancestry data for the birds, and the quality of the birds is just as variable. You can get bad birds from good birds, and less likely, good birds from bad. In either case you may well end up with traits you weren't expecting. Out-breeding tends to lead to heterozygous traits (where in-breeding tends towards homozygous traits), and can also give rise to the phenomenon of heterosis, where the offspring is better than the parents, hence why a breeder with two good lines may attempt to cross them in the hope of getting even better offspring.
As would be expected, a breeder ends up with their own line by line-breeding their birds. How long it takes is not something that is set in stone and varies from breeder to breeder, but it does take quite a number of generations.
Copyright Israel 2013. Used with permission.
Talking to Tim
A look at the Dunedin Botanic Gardens Aviaries
By Israel
The Dunedin Botanic Gardens, like many council run gardens around the country, is home to a fabulous collection of aviaries. The thing that makes this one special, besides its size, range, and health of birds, is that the Bird Keeper, Tim Cotter, is also a member of the Dunedin Bird Club.

|
|
These aviaries, which are mostly occupied by Kaka, were the last ones built. A lot of the funding for them came from sponsors, for an aviary for natives. |
So we thought it would be interesting to go along and see him and have a chat about the aviaries and the birds, and put together a report for the club website and newsletter, showcasing what is available for the public to view, and a little bit of background on it all. This is, of course, that report.
The Dunedin Public gardens is celebrating its 150th year jubilee this year (having been founded in 1863), but interestingly enough some of the aviaries are the oldest part of the gardens, at about 147 years old. The aviaries used to belong to the acclimatization society (who among other things introduced all the foreign birds to NZ) and in the aviaries they kept a number of the birds from their homelands such as Robins, Blackbirds, Thrushes, and their ilk, for public viewing. At that time the gardens were situated on a site by the Leith River (which is now occupied by the university) but it moved to its current location in 1869 due to extensive flooding in 1868.

|
| A Scarlet Macaw. |
Tim, who has both a zoology degree and a vet nursing certificate, has been full-time Bird Keeper for the Gardens aviaries for 8 years, part time for 3 years before that, with his predecessor having held the position for 27 years. There are over 50 species of birds that he looks after, with the help of a part timer, from the tiny Orange-Breasted Waxbill (measuring ~9cm) to the significantly larger cockatoos and macaws (who measure around 81cm), and including both foreign and native birds (such as the Kea and Kaka). While the most popular birds at the aviaries for visitors are the talking Cockatoos or the brightly coloured birds (or the natives if they are tourists who have seen all the others elsewhere), Tims favourite bird at the aviary is the Kaka. Yes, they appear to be a plain parrot, but they have personalities and do learn (unlike the Kea who have a one tracked mind). The Gardens is involved in a breeding program for the South Island Kaka, which is also the rarest native in the aviaries with the population sitting around 5000 birds in the wild (most of which are males). Some of the offspring from the breeding program are released into the wild at the Orokonui Eco Sanctuary. Given their diet (of fruits, berries, seeds, flowers, invertebrates, nectar, sap and so on, with Huhu grubs being a particular favourite which the digs out) and their tendency to chew everything, the Kaka are probably the hardest of the birds to look after (especially since they also have to breed them as well which requires extra attention), while birds like the Bronze Crested Pigeons are the easiest, as they can pretty much be left alone to do their thing. The rarest foreign bird at the aviary is the black-hooded red siskin finch (which was bred with canaries to produce the red-factor canary. This may be part of the reason why their wild population decreased so much as the demand for these vivid birds soared one the success was discovered.

|
| One of the Kaka at the Gardens aviaries. The colour of their plumage was designed to blend in with their surroundings and hide them from their natural predators. Unfortunately it doesn't do much to protect them from the introduced predators who don't rely as much on sight when hunting. |

|
| The black-hooded red siskin. |
Given the limited space at the gardens aviaries there are many considerations that they have to factor in when deciding what birds are allowed to breed (besides the Kaka of course). Importantly, most of the offspring that won't be kept at the gardens have to have a destination already in mind, such as exchanges with zoos, aviaries at other gardens (such as the aviary at Queens Park in Invercargill), or aviaries of breeders who have put in a request for a particular breed of bird. As a result, many of the birds at the aviaries are just of the one sex (such as the siskins which are all males) with the other sex only brought in if needed.
Some of the birds at the aviaries can live for a very long time, making the gardens aviaries a suitable home for them. The longest lived bird at the aviaries was a 130yr old sulphur crested cockatoo (who on average live upwards of 70 years with 100 years not being unknown) and the oldest is Sid, also a sulphur crested cockatoo who was born in the aviaries of the 17th December 1987 (making him 25 at the time of writing). Cockatoos bond with their owners when kept in captivity, and it can be very jarring for them when their owner passes on or they are transferred to new owners. In this setup it isn't as much of an issue and they are guaranteed to be looked after their whole lives. This may be part of the reason why they aren't being breed here at the moment.

|
| Sid the Sulphur Crested Cockatoo. He loves attention and is more than happy to talk to any visitors to his aviary. |

|
| Another block of the aviaries. |
Special attention does need to be paid to some of the species kept, in order for them to do well in their environment. As mentioned, Kaka have special diets, as do Eclectus Parrots (for which the male and female look completely different, so much so that they were originally thought to be two different species) who have a diet somewhat akin to Lorikeets, being soft food, live food (grubs, insects etc) and vegetables. Gauldian finches also need monitoring as their yearly moult is effect by the previous year. Artificial lighting is provided for the finch species during winter (little lights above their feeding dishes) so that they can feed when needed and don't have to spend our long winter nights hungry, and then spend all day out of view eating. Some of the tropical birds have special heaters in the back of their aviaries as well that trigger when frosts hit, to take the chill of the air and keep them warm if needed. They have shown the tendency however to display themselves proudly against the snow seemingly uncaring of the temperature.

|
|
One of the resident Kea, doing a good job of blending in with the background. |
Interestingly, DOC has a Kea Captive Management Plan that requires a certain number of Kea to be kept in captivity. This plan aims to establish a self sustaining and genetically diverse population of Kea in captivity which will support the conservation of the species in the wild.
Many of the aviaries have plants in them now. They are all cultivated in the gardens nurseries, and are selected such that the birds don't kill them and to provide shelter for the occupants. NZ natives in particular like to shade during the warmest parts of the days, while many of the foreign birds take the opportunity to bask in the sun. And for those birds that like live food, the plants also attract their fair share of insects.
From our experience, the Botanic Gardens Aviaries are amongst the best that New Zealand has to offer, and is well worth a visit by any bird fancier, or any one who has a passing interest in our feathered friends. The Dunedin Botanic Gardens was awarded the status of Garden of International Significance by the NZ Gardens Trust in 2010, with the only other one in the South Island being Larnach Castle.
The botanic gardens aviaries consists of 4 clusters of aviaries, with a total of 29 Enclosures. They are of varying sizes, suited to the variety of birds that they hold.
Budgie Genetics
By Israel
When breeding any type of creature it is useful to have an idea what the offspring will be, given a certain pairing of parents, so that appropriate decisions can be made to get the desired results. Many breeders go about this via trial and error, but with a basic understanding of genetics, and what mutations exist for the creature in question, it becomes much easier to breed the varieties that are desired. This of course relies on good knowledge of the make-up of the parents as well.

|
| Budgies of many different colours & varieties. |
This article will look at the genetics of budgies, and is a follow on from the Colour Canary Genetics article published earlier, and hence assumes a basic understanding of the different types of genes. To quickly recap however:
Traits can be of the following types:
- Dominant - trait shows if gene is present in either pair (eg RR or Rr); if both the dominant gene and recessive gene are present they are said to be split for the recessive trait.
- Recessive - trait shows only if it is the only one present (eg rr)
- Semi Dominant - shown phenotype is a combination of the two present genes (eg RY)
- Co-dominant - where both traits show up in the phenotype
- Sex-Linked Recessive - recessive trait that is carried on the sex chromosome (eg ZaW or ZaZa). Hence hens cannot be split for sex-linked traits while males can.
Traits can also have a flow/order of dominance as well, and results of pairing are found by using punnet squares, which give a percentage likelihood for each chick in a clutch.
Colour mutations - the six basic colours

|
|
Lacewing Yellow hen. The yellow base colour can be clearly seen. |
Like many birds budgies have two colour layers to their feathers - they have the base colour / pigmentation, and then they have the melanin pigmentation (the type in budgies is known as eumelanin, and the way it reflects light causes it to appear as a blue colour) on top of that. The combination of these two pigments is what gives the budgies their colouring. The eumelanin pigmentation does not cover the entire body however, in particular it is not present on the mask feathers of budgies, and on the wings it provides the black colour that give them their patterns.
Budgies have two base colours (a pigment base), Yellow and White, with the former being dominant to the latter. The symbol B is usually used to represent base colour in budgies, so B is Yellow, and b is white. With the exception of the yellow-faced mutation, the base colour can be seen in the mask feathers of the budgie. The common green budgie results from the yellow base colour mixing with the blue colour layer in the body feathers (eumelanin), resulting in a green bird. When the base colour is white the blue feathers result in a blue bird.
| Dominant | Recessive | |
| Base Colour | Yellow (B) | White (b) |
As would have been noticed there are three different variations to the green colour in budgies, which are typically termed green/light-green, dark-green, and olive. This is because of the 'dark-factor' trait, which effects the darkness of the eumelanin pigmentation in the body feathers. The dark factor gene, represented by D (and lack by d) is semi-dominant, which is why we end up with the three grades of colour. All budgies possess both the dark-factor gene and the base colour gene, and in fact either the presence or absence of all the other mutations as well, as is the case for all species. But when doing any calculations for budgies both of these traits should be involved. In cases where the dark-factor pigmentation does not show (for example clear birds) the bird will still possess the factor, but its nature cannot be visually discerned. The intensity of the dark factor can vary a little as well, so not all olives will be the exact same colour for example.
| Base Colour / Dark-factor | absent (dd) | single (Dd) | double (DD) |
| Yellow | green | dark-green | olive |
| White | skyblue | cobalt | mauve |

|
|
Lacewing White hen. The white base colour can be clearly seen. |
Yellow-based birds are often called the green-series, while white-based the blue series.
So, lets look at an example. Lets pair a dark-green budgie with a sky-blue hen. The hens genotype will be bbdd, but there are two possible combinations for the dark-green male, his base colour yellow, which is dominant. He will either be homozygous yellow, or heterozygous and split for white. For the sake of variety let's assume that he is split for white, so that his genotype is BbDd. The punnet square will look as follows, removing any duplicate lines (the hens only gene combination is bd, so to avoid duplicating it four times it will be listed just once):
| BD | Bd | bD | bd | |
| bd | BbDd | Bbdd | bbDd | bbdd |
The resulting offspring will be:
- 25% dark-green split for white (BbDd) (or single factor dark-green)
- 25% green split for white (Bbdd) (or single factor green)
- 25% cobalt (bbDd)
- 25% sky-blue (bbdd)
Colour Mutations - adding more variety

|
| Normal Grey-factor male. |
There are a number of mutations which add additional colour to budgies feathers. The Grey-Factor, which as its name suggests adds a grey colour to the feathers, is a dominant trait. It is represented by G, while its absence is represented by g. In a yellow-based bird their colour becomes grey-green, while a white-based bird becomes grey. It effects both the eumelanin (quite significantly, mostly overriding the original colour) and the base colour (slightly- usually dulling the colour of the mask). The dark-factor is muted by the grey-factor.
The Violet-factor is a semi-dominant colour adding trait (represented by V), however it isn't very strong and usually only shows up noticeably on sky-blue (when present as VV) or cobalt budgies (VV or Vv), for which the blue colour of the feathers becomes a violet colour. In the other colours, especially for yellow based birds, as a double factor its effect is similar to the dark-factor. Sky-blue with the single factor may have a violet tinge to their feathers.
The Slate mutation is a rare sex-linked mutation that darkens the blue colour in blue series birds, and acts like the darkening-factor in green series.
| mutation | notation | dominance |
| Grey Factor | G/g | dominant |
| Violet Factor | V/v | Semi-dominant (only shows up in sky-blue/cobalt's) |
| Slate | Zl | Sex-linked recessive |
Dilutions
Budgies have three different types of dilutions (as well as the absence of any dilution which becomes the fourth type) which all share the same position on the chromosome (so you will always end up with a combination of two of the four different possibilities on a bird). The dilutions are all recessive to normal (C). The three types of dilution are dilute (cd) which is recessive to the other three, and greywing (cg) and clearwing (cw) which are co-dominant to each other. Hence the dominance flow for the dilutions looks like this:
Normal > greywing = clearwing > dilute
For some clarification, in order for dilution to show in any form both genes must carry one of the three dilutes genes, otherwise they birds phenotype will be normal. In order for dilute to show both genes must be dilute. If either of the genes is greywing or clearwing with dilute, then that trait will show instead of dilute. If both greywing and clearwing are present then both show (and the bird is said to be full-body-colour greywing).
In greywings the body colour is diluted by 50%, and the black melanin on the wings etc become grey. In clearwings the black melanin becomes almost entirely muted but the body colour is brightened instead. When combined for full-body-colour greywings the black melanin is turned grey but the body colour is brightened. Dilute budgies are diluted all over, and both the body colour and black melanin are very light.
Looking at an example of crossing a normal budgie split for greywing (Ccg) and a full-body-colour greywing (cwcg):
| C | cg | |
| cw | Ccw | cwcg |
| cg | Ccg | cgcg |
The resulting offspring would be:
- 25% Normal split clearwing (Ccw)
- 25% Normal split greywing (Ccg)
- 25% Full-body-colour greywing (cgcw)
- 25% Greywing (cgcg)
| mutation | notation | dominance |
| Greywing | cg | semi-dominant/recessive |
| Clearwing | cw | semi-dominant/recessive |
| Full-body-colour Greywing | cgcw | |
| Dilute | cd | recessive |
Other Traits

|
| Opaline Cobalt Male Budgie. |
The Ino mutation is a sex-linked recessive mutation removes all pigmentation and markings apart from the base colour and gives the bird red eyes. As is typical the Ino mutation masks almost all other mutations. However it only partially masks the cinnamon mutation (which becomes known as the lacewing mutation when combined with Ino). It also doesn't completely suppress the yellow face mutations.
The Opaline mutation is also sex-linked recessive, and reduces the intensity/size of the stripes down the back of the budgies neck (so that there is very little black), and causes the body feather colour to run through the wing feathers of the bird as well (as opposed to the base colour).
The Spangle mutation is a dominant trait, and the single factor version of the mutation causes the markings on the tail and wing to be inverted (so mostly base colour (or series colour if combined with the opaline mutation) with thin black lines at the edge). Present in a double factor the bird is either clear yellow or clear white, depending on its series. However, unlike the Ino's they don't have the red eyes.
As is with other birds (eg Canaries and Zebra Finches) the Cinnamon mutation causes the black melanin to turn brown, giving the budgies brown markings on their wings and tails. It can also give a brown tinge to their body colour as well. The cinnamon factor is sex-linked recessive.
There are three forms of the Pied mutation on budgies - recessive, dominant and clearflight pied. The dominant pied mutation as a single factor causes a few clear (base-colour) patches around the belly and bottom of the wing. Double factor dominant pieds lose most of their markings. Recessive Pieds also lose most of their markings, but their dark-plum eyes do not lighten with age as they do for the dominant pieds. Clearflight pieds lose their markings only on their flight feathers and tail feathers. Crossing a clearflight and recessive pied gives rise to the dark-eyed clear (PPrr or Pprr) variety, for which the melanin is completely suppressed (ie it is a clear bird). Unlike the Ino's however, the Dark-eyed Clear has, as its name suggests, dark-plum eyes.
| mutation | symbol |
|
dominance | ||
| Ino | Zi |
|
sex-linked recessive | ||
| Cinnamon | Zc | sex-linked recessive | |||
| Ino + Cinnamon | Zci | Lacewing | sex-linked recessive | ||
| Opaline | Zo | sex-linked recessive | |||
| Spangle | S/s | dominant | |||
| Dominant Pied | T/t | (semi)-dominant | |||
| Recessive Pied | R/r | recessive | |||
| Clearflight Pied | P/p | dominant |

|
| Spangle Light-Green Male Budgie. |
As an example, lets pair a sky-blue cock split for Ino with a single factor green-cinnamon hen (ie she is split for white). Their genotypes respectively are bbddZZi and BbddZW. Since they are both absent the dark-factor it can be removed from the punnet calculations. The male also has only 2 combinations bZ and bZi, while the hen has four.
| bZ | bZi | |
| BZ | BbZZ | BbZZi |
| BW | BbZW | BbZiW |
| bZ | bbZZ | bbZZi |
| bW | bbZW | bbZiW |
The resulting offspring would be:
- 12.5% single factor green male (BbZZdd)
- 12.5% single factor green hen (BbZWdd)
- 12.5% sky-blue male (bbZZdd)
- 12.5% sky-blue hen (bbZWdd)
- 12.5% single factor green male split for cinnamon (BbZZidd)
- 12.5% single factor cinnamon-green hen (BbZiWdd)
- 12.5% sky-blue male split for cinnamon (bbZZidd)
- 12.5% cinnamon sky-blue hen (bbZiWdd)
Complicated Traits
Yellow-face I & II are somewhat complicated mutations that are not fully understood and rely on multiple genes. At this stage they are beyond the scope of this article for modelling, but we shall have a brief look at them.
Yellow-face I, in its most basic modelling, is considered to be recessive to the wild-type (ie like the white-base colour aka blue series), however it only shows up in the phenotype when it is single factor in the blue-series. Single factor green series look just like greens, and double factor yellow-face I's look just like sky blues. Single factor blue series Yellow-face I's look like blues series, except that the forehead and mask are yellow instead of white.
Yellow-face II is also considered recessive to the wild type. The mutation does not show up in single factor green series. Double factor yellow-face II's look similar to single factor blue-series Yellow face I's except that the yellow colour is brighter and it leaks into the body feathers a little as well. Blue series single factor yellow-face II's have the yellow mask and forehead, and the yellow is diffused throughout the body feathers after the first moult, giving them a colour that is between a green and a blue.
There are many more mutations for budgies (around 30 according to wikipedia), but as they are not present in New Zealand they will not be covered by this article. Many of the above mutations can be combined as well to give all sorts of interesting results and varieties. As with all birds, good recording is essential to success when breeding, especially when trying to breed for certain colours or traits.
Bibliography:Wikipedia entry on budgie colour genetics
Wikipedia entry on the yellow face I mutation
Wikipedia entry on the yellow face II mutation
Budgie Place page on budgie colours
Copyright Israel 2013. Used with permission.
What to put in?
A look at soft foods from a canary & budgie perspective
By Israel
Soft foods, also known as Nesting Foods or Rearing Foods, are an important part of a cage or aviary birds diet, in particular during breeding and moulting season, but also year round. The basis of many caged bird diets is seed, but by itself seed isn't nutritionally complete. It needs to be complemented, at a minimum, by some fresh vegetables and fruit. During breeding and moulting season the protein requirements of a birds diet goes up quite considerably and a basic diet will not be able to provide enough protein to maintain top quality birds or raise prime quality chicks. This is where soft foods come in, as a source of extra protein for the birds, as well as any vitamins, minerals, and whatever else the breeder thinks may be lacking or needed in the birds diet. You will find that each breeder is likely to have their own special recipe that they use for their birds, and reasons for why they do so. It is a reflection of individual views, knowledge, and learning, as well as how much money and time a breeder is willing to spend on the various components that make up a particular mix. Success will vary, and what works well for one person may not for another.

|
|
Softfood mix containing Egg, Rape Seed & Breadcrumbs. |
Soft food mixtures come in many different levels of complexity and composition, with the complexity seeming to have risen over the last half a century compared to what our grandparents used. "Back in the day" the mixes were as simple as crushed wine biscuits and milk, or mashed boiled egg and breadcrumbs (of which the latter has been successfully used by the author). Now days the mixtures can include a dozen or more different ingredients.
When putting together a soft food recipe there are two questions that should be asked of each ingredient - why am I adding it and to a lesser extent how much, as some ingredients are not very beneficial or economical in large quantities. The main purpose of soft foods is to provide an (additional) source of protein, which is necessary for chick development and feather growth*. Secondary reasons include providing missing or extra vitamins and minerals, in particular calcium, and extra carbohydrates beyond what seed provides. Variety is another purpose, as like humans birds can get somewhat bored eating the same food all the time. Many vegetables (and vitamins and minerals) also help improve feather quality and health. Finally taste can be a factor in acceptance of various foods, so ingredients can be added to effect the overall taste or offset another ingredient. All ingredients should be considered against these factors.
The components of soft foods can be separated into three categories - dry, wet, and extras. The dry form the bulk of the soft food mix and large quantities can be prepared before hand (being broken down and mixed together). The wet ingredients which make up a smaller portion of the mix need to be prepared and added on the day that it is served, with the extras usually being done on a case by base basis.
Dry components:- Breadcrumbs are the main staple of many commercial soft food mixes (which also usually contain Rape Seed which canaries love), and a source of carbohydrates. Breadcrumbs are nutritionally empty though as a food, being mostly carbohydrates.
- Wine Biscuits such as super wine or milk arrowroot. They are a source of carbohydrates and also sweet, making them more palatable to the birds. Birds can be fussy though, liking one type (or even brand) over others.
- Soy Protein (Full fat Soy Meal being an example) is very high in protein and has found favour with many breeders as a part of their mixture bases.
- Rolled Oats are a good source of carbohydrates. Hulled Oats is often one of the seeds given to budgies, and their rolled form is easier to consume for other types of birds as well as mixing better into a soft food.
- Weetbix is a source of protein, carbohydrates and fibre. Because it is a fortified cereal it also contains some added vitamins and minerals.
-
Wheatgerm is a good source of both protein and carbohydrates, and also high in fibre.
Chick Starter Crumble - Whey Powder has been used by some breeders, again because it is a good source of protein and carbohydrates. Typically a mix designed for human consumption is used, another option being a lactose-free milk powder.
- Chick Starter Crumbs**, of which there are a number of varieties, are a premade mix that contain many of the required ingredients of a complete rearing food (protein etc), and are used as a part of soft food mixes by a number of the society members.
- Cornmeal / Ground Maize is another bulking ingredient in soft foods, and is a good source of carbohydrates.
- Mashed Boiled Egg is high in protein and amino acids and is considered by some to be a complete food (it is for the baby chickens that grow up in the eggs at least).
- Vegetables and fruits are sources of vitamins and minerals, moisture, fibre, and can promote good feather quality and colour. They can be feed outside of a soft food mix as well, and common ones are Carrots, Broccoli, Celery, Silverbeet, Sweet Potato, Red Peppers, Beetroot, Apple, Corn, Cucumber, and Pumpkin. Fruit and vegetables should be grated or blitzed in a food processor.
- Soaked Rice is a good source of calcium and carbohydrates. Rice should be soaked for 24 hours. It can also be cooked in a microwave beforehand.
- Soaked Seed is also a good source of vitamins and minerals and is readily consumed by birds. Care needs to be taken that the seeds do not go mouldy though.
- Spirulina is a source of protein, minerals, vitamins, and amino acids. Only a small amount should be used as it is very concentrated and excessive amounts can cause digestive issues.
- Ground / Crushed Dried Kelp is also a source of vitamins and minerals, and again is used in small quantities.
- Liquid Vitamin Supplements are also added by some breeders, either to a soft food mix or to the drinking water.
- Crushed Egg Shell or Oyster Grit is sometimes added as an extra source of calcium. Egg shells must be baked first to kill any bacteria left on the shells.
What ingredients are used, or even if a soft food mix is used in the first place, comes down to the individual choices of a breeder. As mentioned already success will vary from one breeder to another with any particular mix, or even from year to year for the one breeder. There are many reasons, amongst these the full diet of the birds, their environment, and even the birds themselves. Some birds are just plain fussy! Familiarity is also an important factor, as birds aren't always willing to readily accept a new food, and hence it is inadvisable to make significant changes to soft food mixes during breeding season as it is likely it will be rejected and you'll loose chicks as a result. Feeding soft food year round (in smaller quantities outside of the breeding and moulting season) is a good way to maintain the birds familiarity with it and promote good health.
Finally, when buying new birds it is always worth asking what diet they have had thus far. For those new to the hobby it will provide a good basis (you'll even be able to keep them on the same diet), and for the more experienced breeders it can aid in decision making and may help identify issues that arise later.
* The amount of protein required for optimal growth isn't fixed in stone and will vary from one publication to another. The general indication from information viewed seems to put the recommended percentage of protein of a birds full diet to be around 20-25%.
** There are two medicated (with a coccidiostat) brands of chick starter crumble currently available - Reliance & NRM.
Copyright Israel 2013. Used with permission.
A Nest is just a Nest?
By Israel

|
| Nest of a wild bird, likely a wax-eye in this case. |
The kingdom of birds if filled with incredible variety, and not just with the type, shape, and feather colour of birds, but also in the wide variety of nests that birds do (or in some cases don't) build. To the average person, when you mention a nest they think of a half cup style nest woven out of twigs and whatever else the bird could find, and while many birds do use that 'template' style, it is by no means the only one, and again, while some birds nest in trees, others like the native Kiwi or penguins, are ground nesters. Granted that both of those species are flightless. Of course there are also parasitic nesters - such as the Cuckoo, who lay their eggs in the nests of other birds, and leave them to do all the hard work of raising the chicks.
Even amongst the caged & aviary birds that we keep there is much variety, and this means that we do need to pay attention to the needs and habits of our birds in order to lay the groundwork for successful breeding.
Canaries follow the basic nest template, creating woven nests out of whatever material they can lay their beaks on - be it think twigs, grass, threads, twine, shredded newspaper or the like. However, they don't build them free-standing but prefer to build them in an open top nesting dish or other similar container, be it seed dish, yoghurt pottle, or other concave dish. Usually they also prefer to nest high, but they have been known to build them on the floor in some cases. To give them a label for their nest building style, canaries are weavers. [As a note some canaries are lazy and will lay eggs in a nesting dish that just has a felt nest liner and nothing else, but that is more a characteristic of the bird itself than the species.] Blackbirds are another example of weavers, and they build their nests with plant material low down in dense shrubs, trees, climbers and so on, assuming there is adequate cover; occasionally they will build them on the ground too.

|

|
|
Canary nest built in a seed dish, with 5 red-factor chicks nestled inside. |
Canary nest built in a wicker nesting basket |
Zebra finches prefer enclosed nests, with all sides closed in apart from a small hole at the front. When the provided space doesn't quite meet their needs, assuming that they take to it, they will build the nest up so that it does cover them completely leaving a small entry hole. The zebra finch style of nest building can be described as packing - they pack the provided nesting material inside the nest and compact it down, but without the weaving finesse of some of the other birds. They are also prolific nest builders and will build layer on top of layer inside the nest, even on top of un-hatched eggs, until they run out of space. Bengalese finchs are like Zebra finches nest wise, but rather than one pair per nest many pairs will try and cram into the same nest if able (as per their name - Society Finch). Swallows are another example of packers, however they build their nests from scratch using small balls of mud which they pack together to form an enclosed space. Often swallow nests can be found under the eaves of buildings near water.

|

|
|
Wicker nesting basket designed for finches, with a zebra finch chick sitting on the top |
Enclosed nesting box designed for finches. |
Budgies also prefer enclosed nests, but unlike canaries and finches they don't bother with building up a nest inside, instead laying their eggs on the nesting box floor and hoping for the best. Breeders often put sawdust or woodchips inside the nesting box, but as often as not these are kicked to the side with the budgies laying their eggs in the middle (or off to the side if the box is too big). As this can lead to eggs rolling off to one side and being ignored, some breeders have taken to making a concave shallow in the floor of the nesting box so that the eggs stay under the bird. Cockateils have similar nesting habits, with their nesting box being an just an enclosed space in which they lay their eggs. Again some breeders line their nests with wood-chips or even paper towels to absorb some of the moisture.

|

|
|
'Back' side of a budgie nesting box. The front side is similar to the finch nesting box above. |
Inside of a budgie nesting box, which has been lined with bark, with three chicks and two infertile eggs. |
As with any creature, birds all have individual personalities, and this will effect their nesting habits and preferences. Keeping records of these details will be invaluable to the breeder and contribute to success, or at least to keeping the birds happy :)
Copyright Israel 2013. Used with permission.